Гаплогруппы евреев. Генетика евреев
Часть 2. Городские легенды о ДНК-генеалогии
2.1. Гаплогруппа J1 – еврейская?
– Слушай, я тут результаты получил. У меня гаплогруппа J1. Народ с форума говорит, что J1 полно у евреев. Еврейская, типа, гаплогруппа. Так что, я «из колена Авраамова» что ли?
– Ты губищи-то не раскатывай! «Еврей», видите ли, он!
– А кого тогда ДНК-генеалогия называет потомками Авраама? Разве не тех, кто из гаплогруппы J1?
– Да, но не всех подряд, а только тех, кто относится к субкладу J1c3 и имеет гаплотип типа 12 23 14 10 13 17 11 16 11 13 11 31
– Тогда кто такие остальные J1, которые не J1c3?
– Да кто угодно. Гаплогруппа появилась аж целых 20 тыс. лет назад. Можешь себе представить, сколько людей наплодилось с того времени и сколько народов они успели сформировать!
2.2. Если русский, то только R1a1?
– У меня к вам очень важный вопрос. Я посмотрел множество статей по гаплогруппам русских. Основной является R1a1.
– Если быть совсем точным, то не только R1a1, но и I2a и N1c1. Но в чём суть твоей проблемы?
– Проблема в том, что моя гаплогруппа – T. Она, в отличие от перечисленных вами гаплогрупп, никак не могла быть у восточных славян. Значит, кто-то из предков не был славянином и относительно недавно влился в русский народ. У меня появились сомнения – можно ли считать меня русским, если я не принадлежу к гаплогруппам большинства русских?
– Вопрос ясен. Теперь давай выясним, является ли русским носитель R1a1 индус Джавахарлал Кумар? Ответь мне на вопрос: Кумар или его предки когда либо жили в стране под названием Русь или Россия? Не на древней территории, которая 5000 лет спустя стала называться Русью, а именно на Руси или в России?
– При такой постановке вопроса ответ прост – нет, не жили.
– Он сам или его предки когда-либо владели русским языком?
– Нет, не владели.
– У него европейская внешность?
– Нет, он южноазиат.
– Он или его предки справляли масленицу? Есть ли в их традиции запрет здороваться через порог или встречать гостя хлебом-солью?
– Нет, всего этого у него нет.
– Его дед, прадед или прапрадед были православными?
– Нет, не были.
– Его предки воевали с Наполеоном в Бородино?
– Нет, не воевали.
– Ну вот, а теперь подумай, может ли этот человек только на основании одной гаплогруппы R1a1 считаться русским?
– Да, вроде всё так, но…
– Что, неубедительно всё это? Тогда просто подумай, к кому обращены воображаемые строки Невского «Вставайте люди русские!» Неужели только к тем, у кого R1a1, включая индусов, шотландцев, казахов, киргизов? Нет. Эти строки касаются лишь тех, к кому применимо всё то, что не применимо к досточтимому Кумару. Даже если у этих людей не R1a1, I2a2 или N1c1, а H, L или T.
2.3. Как тестируют целые народы?
– Слушай, у меня N1c1, посмотрел по интернетам, пишут, что максимум N1c1 – у удмуртов. Аж целых 75%! Что-то мне не верится, что кто-то обошёл и протестировал лично все 500 тысяч удмуртов!
– Как по-твоему, Y-гаплогруппа у кого бывает?
– Ну, только у мужиков, конечно!
– Сколько всего на свете удмуртских мужиков?
– Всего удмуртов 500 тысяч, из них половина – мужики, получается 250 тысяч человек. Но всё равно, даже 250 тысяч человек проверить нереально! НЕ-ВЕ-РЮ!
– Как ты думаешь, какая гаплогруппа у твоего отца?
– Разумеется, такая же, как у меня – N1c1. Точнее, у меня такая же, как у него!
– А у твоего брата какая?
– Само собой тоже N1c1, что за глупые вопросы?
– У твоего отца есть брат? У тебя есть родной дядя по отцу?
– Есть, но какое это имеет отношение?
– Какая у дяди может быть гаплогруппа?
– Блин, чего спрашивать-то и так же ясно, что N1c1!!!
– А у сыновей твоего дяди, твоих двоюродных братьев, какая гаплогруппа?
– Да та же самая, что и у меня!
– Твой дед по отцу жив?
– Да!
– У этого деда какая гаплогруппа может быть?
– N1c1 конечно же! И у его брата тоже N1c1, и у его сыновей тоже. У всех мужиков из моего рода будет N1c1!
– Теперь ты понял, что для выяснения гаплогруппы всего твоего рода с отцом, братьями, дядьями и всего прямого мужского кагала достаточно протестировать тебя одного?
– Кажется, начинаю понимать! Значит, чтобы проверить целый народ, надо просто собрать только тех мужчин, которые не считают друг друга родственниками, то есть представителей разных родов?!
– Правильно, молодец! Ты видишь, что у одного человека в среднем есть около 30 живых родственников по прямой мужской линии. Просто не всех он знает лично, а может, и не видел никогда. А если брать наших дедов, так у них семьи по 4-5 сыновей совсем не редкость.
– Да, могу себе представить, сколько у моего прадеда может быть потомков вместе с дедом, его братьями и их внуками! Действительно, человек 30, а то и больше!
– Соображаешь! А теперь прикинь, что из 250 тысяч удмуртов на самом деле можно тестировать как минимум в 30 раз меньшее число людей!
– Но всё равно, даже если 250 тысяч разделить на 30 получится что-то в районе 8000 человек. Всё равно много!
– На самом деле, чтобы вычислить набор гаплогрупп такого по численности народа как удмурты, надо не 8000 человек, а ещё меньше.
– Это почему?
– Да потому что из этих 8000 человек где-то каждые человек 50 также представляют собой род. Просто эти люди этого не знают или не помнят. Так часто бывает, особенно если люди родом из одного региона. Их общий предок жил в чуть более отдалённом прошлом, данных по нему не осталось и поэтому они и не сознают своего родства.
– То есть вы хотите сказать, что для тестирования удмуртов достаточно было обследовать 160 человек из разных областей?
– Да, примерно так.
– Но ведь для этого надо колесить по всей Удмуртии!
– Нет, не обязательно. Достаточно прийти в какой-нибудь ВУЗ в том же Ижевске и выбрать несколько студенческих групп. Там как раз собраны удмурты со всей Удмуртии. Надо лишь перед тестом удостовериться, что среди проверяемых нет родственников.
– Да, действительно, дело упрощается.
– Так ты сможешь, не выходя из Ижевска, составить более-менее правдоподобную картину
о наборе гаплогрупп удмуртского народа.
– А почему «правдоподобную»
, а не, скажем, полную
?
– Потому что всегда есть вероятность того, что в твою выборку (то есть в группу людей, которую ты тестируешь) не попали представители каких-нибудь гаплогрупп, относительно редко встречающихся у данного народа. Но выявить, какая гаплогруппа наиболее характерна интересующему тебя народу, ты всегда сможешь.
– Да, теперь я понял. То есть если я вижу информацию, что у удмуртов 75% N1c1, то это означает, что у любого произвольно взятого удмурта с вероятностью 75% может быть выявлена гаплогруппа N1c1. Логично.
В Испанию , Пакистан и другие регионы.
Субклады
Известные субклады J1 (M267) и соответствующие им SNP мутации по ISOGG-2010:
- J1 M267
- J1* -
- J1a M62
- J1b M365
- J1c L136
- J1c* -
- J1c1 M390
- J1c2 P56
- J1c3 P58
- J1c3* -
- J1c3a M367, M368
- J1c3b M369
- J1c3c L92, L93
- J1c3d L147
- J1c3d* -
- J1c3d1 L222
- J1c3d1* -
- J1c3d1a L65.2/S159.2
- J1c3d1* -
- J1 M267
Даже по результатам раннего коммерческого тестирования сложилось впечатление, что большинство современных носителей данной гаплогруппы относятся именно к подгруппе J1c3 - ветвь P58 , что уже ныне подтвердилось более широким тестированием и является очевидным фактом.
Распространение
Ближний Восток
Самая многочисленная подгруппа J1c3 (P58) распространёна в значительной степени среди евреев и населения Аравийского полуострова . Так же гаплогруппа в целом широко распространена в Леванте и среди семитоязычного населения Северной Африки , например ассирийцев, палестинцев-христиан, друзов. Для евреев характерны субклады J1c3* и J1c3d* (коэны). Например корневой субклад J1* (M267), будучи характерным для ассирийцев , народов Дагестана , Чечни , европейцев, у евреев и арабов практически не наблюдается. Наибольшая концентрация данной гаплогруппы, субклад J1c3d* , наблюдается в Йемене (76 %) и Саудовской Аравии (64 %) , среди палестинцев (38 %) , в Сирии (30 %) и Ливане (25 %) .
В Восточной Анатолии значительно представлен субклад Z1842.
Частота гаплогруппы J1 заметно падает на границах арабских стран и Дагестана с другими странами, такими как Иран (10,5 %) и Турция (9 %) .
Северная Африка
Частота J1 в Северной Африке: Алжир (22 %), Египет (21 %), Тунис (17 %), Марокко (7 %).
Кавказ
В регионе Кавказа наиболее распространён макросубклад Z1842. Высокая концентрация этой гаплогруппы, особенно указанного Z1842, наблюдается среди даргинцев (84 %), аварцев (67 %), лезгин (44 %), чеченцев (35 %), кумыков (21 %).
Европа
В целом частота J1 в Европе не высока. Однако относительно высокие частоты были зафиксированы в центральных Адриатических районах Италии Горгано (англ. ) (17,2 %), Пескара (15 %), Паола (11,1 %), южносицилийской Рагузе (10,7 %) , на Мальте (8 %), Кипре (6 %) . Европейские субклады принадлежат к древнему неолитическому населению Европы, и значительно отличаются от арабских и еврейских субкладов по временным дистанциям до общего предка В Наиболее представленным в Европе, в том числе и Восточной, является субклад L1128 и его подветвь L1189, хотя на Европейском театре ДНК исследований периодически наблюдаются и макросубклад Z1842, который сам по себе преимущественно распространен в Восточной Анатолии и на Кавказе. Обнаружена исключительно восточно-европейская подветвь L1189, характерная для славянских народов.
Известные представители гаплогруппы J1
- Армянские князья Мелик-Шахназаряны ; макросубкалад Z1842.
- Дворянский шотландский род Грэмов - маркизы и герцоги Монтроуз; макросубклад P58.
Напишите отзыв о статье "Гаплогруппа J1 (Y-ДНК)"
Примечания
- Semino et al. 2004"
- Arburto et al. 2008
- Alshamaly et al. 2009: 84/104 (81 %), Malouf et al. 2008: 28/40 = 70 % J1-M267, 6/40 = 15 % J2a4b-M67
- Cadenas AM, Zhivotovsky LA, Cavalli-Sforza LL, Underhill PA, Herrera RJ (March 2008). «Y-chromosome diversity characterizes the Gulf of Oman». Eur. J. Hum. Genet.
16
(3): 374–86. DOI :10.1038/sj.ejhg.5201934 . PMID 17928816 .
Yemen 45/62 = 72,6 % J1-M267
Qatar 42/72 = 58,3 % J1-M267 - Alshamaly et al. 2009: 68/106 (64 %)
- Semino et al. 2004
- combined (Wells et al. 2001 19 %) & (Zalloua et al. 2008 20 %) Wells et al. 2001: 32,0 % E-M96, 30,0 % J1-M267, 30,0 % J2-M172, 2,0 % L-M20, 6,0 % R1-M173. (Zalloua et al. 2008) 184 out of 914, , Zalloua et al. 2008
- Mean percentage derived from 3/33 = 9,09 % North Iran and 14/117 = 11,97 % South Iran, , Regueiro et al. 2006
- 47 out of 523, , Cinnioglu et al. 2004
- Maciamo. (англ.) . Eupedia. Проверено 4 января 2016.
- Yunusbayev B, Kutuev I, Khusainova R, Guseinov G, Khusnutdinova E (August 2006). «Genetic structure of Dagestan populations: a study of 11 Alu insertion polymorphisms». Hum. Biol. 78 (4): 465–76. DOI :10.1353/hub.2006.0059 . PMID 17278621 .
Ссылки
- Y-DNA_J project of FTDNA
| п Эволюционное древо гаплогрупп Y-хромосомы человека | |||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
|
|||||||||||||||||||||||||||||||
| Y-хромосомный Адам | |||||||||||||||||||||||||||||||
| A00 | A0-T | ||||||||||||||||||||||||||||||
| A1a | A1b | ||||||||||||||||||||||||||||||
| A1b1 | |||||||||||||||||||||||||||||||
| GHIJK | |||||||||||||||||||||||||||||||
| HIJK | |||||||||||||||||||||||||||||||
| IJK | |||||||||||||||||||||||||||||||
Отрывок, характеризующий Гаплогруппа J1 (Y-ДНК)
– Да нечего делать, дружок, – сказал князь, – они все такие, не разженишься. Ты не бойся; никому не скажу; а ты сам знаешь.Он схватил его за руку своею костлявою маленькою кистью, потряс ее, взглянул прямо в лицо сына своими быстрыми глазами, которые, как казалось, насквозь видели человека, и опять засмеялся своим холодным смехом.
Сын вздохнул, признаваясь этим вздохом в том, что отец понял его. Старик, продолжая складывать и печатать письма, с своею привычною быстротой, схватывал и бросал сургуч, печать и бумагу.
– Что делать? Красива! Я всё сделаю. Ты будь покоен, – говорил он отрывисто во время печатания.
Андрей молчал: ему и приятно и неприятно было, что отец понял его. Старик встал и подал письмо сыну.
– Слушай, – сказал он, – о жене не заботься: что возможно сделать, то будет сделано. Теперь слушай: письмо Михайлу Иларионовичу отдай. Я пишу, чтоб он тебя в хорошие места употреблял и долго адъютантом не держал: скверная должность! Скажи ты ему, что я его помню и люблю. Да напиши, как он тебя примет. Коли хорош будет, служи. Николая Андреича Болконского сын из милости служить ни у кого не будет. Ну, теперь поди сюда.
Он говорил такою скороговоркой, что не доканчивал половины слов, но сын привык понимать его. Он подвел сына к бюро, откинул крышку, выдвинул ящик и вынул исписанную его крупным, длинным и сжатым почерком тетрадь.
– Должно быть, мне прежде тебя умереть. Знай, тут мои записки, их государю передать после моей смерти. Теперь здесь – вот ломбардный билет и письмо: это премия тому, кто напишет историю суворовских войн. Переслать в академию. Здесь мои ремарки, после меня читай для себя, найдешь пользу.
Андрей не сказал отцу, что, верно, он проживет еще долго. Он понимал, что этого говорить не нужно.
– Всё исполню, батюшка, – сказал он.
– Ну, теперь прощай! – Он дал поцеловать сыну свою руку и обнял его. – Помни одно, князь Андрей: коли тебя убьют, мне старику больно будет… – Он неожиданно замолчал и вдруг крикливым голосом продолжал: – а коли узнаю, что ты повел себя не как сын Николая Болконского, мне будет… стыдно! – взвизгнул он.
– Этого вы могли бы не говорить мне, батюшка, – улыбаясь, сказал сын.
Старик замолчал.
– Еще я хотел просить вас, – продолжал князь Андрей, – ежели меня убьют и ежели у меня будет сын, не отпускайте его от себя, как я вам вчера говорил, чтоб он вырос у вас… пожалуйста.
– Жене не отдавать? – сказал старик и засмеялся.
Они молча стояли друг против друга. Быстрые глаза старика прямо были устремлены в глаза сына. Что то дрогнуло в нижней части лица старого князя.
– Простились… ступай! – вдруг сказал он. – Ступай! – закричал он сердитым и громким голосом, отворяя дверь кабинета.
– Что такое, что? – спрашивали княгиня и княжна, увидев князя Андрея и на минуту высунувшуюся фигуру кричавшего сердитым голосом старика в белом халате, без парика и в стариковских очках.
Князь Андрей вздохнул и ничего не ответил.
– Ну, – сказал он, обратившись к жене.
И это «ну» звучало холодною насмешкой, как будто он говорил: «теперь проделывайте вы ваши штуки».
– Andre, deja! [Андрей, уже!] – сказала маленькая княгиня, бледнея и со страхом глядя на мужа.
Он обнял ее. Она вскрикнула и без чувств упала на его плечо.
Он осторожно отвел плечо, на котором она лежала, заглянул в ее лицо и бережно посадил ее на кресло.
– Adieu, Marieie, [Прощай, Маша,] – сказал он тихо сестре, поцеловался с нею рука в руку и скорыми шагами вышел из комнаты.
Княгиня лежала в кресле, m lle Бурьен терла ей виски. Княжна Марья, поддерживая невестку, с заплаканными прекрасными глазами, всё еще смотрела в дверь, в которую вышел князь Андрей, и крестила его. Из кабинета слышны были, как выстрелы, часто повторяемые сердитые звуки стариковского сморкания. Только что князь Андрей вышел, дверь кабинета быстро отворилась и выглянула строгая фигура старика в белом халате.
– Уехал? Ну и хорошо! – сказал он, сердито посмотрев на бесчувственную маленькую княгиню, укоризненно покачал головою и захлопнул дверь.
В октябре 1805 года русские войска занимали села и города эрцгерцогства Австрийского, и еще новые полки приходили из России и, отягощая постоем жителей, располагались у крепости Браунау. В Браунау была главная квартира главнокомандующего Кутузова.
11 го октября 1805 года один из только что пришедших к Браунау пехотных полков, ожидая смотра главнокомандующего, стоял в полумиле от города. Несмотря на нерусскую местность и обстановку (фруктовые сады, каменные ограды, черепичные крыши, горы, видневшиеся вдали), на нерусский народ, c любопытством смотревший на солдат, полк имел точно такой же вид, какой имел всякий русский полк, готовившийся к смотру где нибудь в середине России.
С вечера, на последнем переходе, был получен приказ, что главнокомандующий будет смотреть полк на походе. Хотя слова приказа и показались неясны полковому командиру, и возник вопрос, как разуметь слова приказа: в походной форме или нет? в совете батальонных командиров было решено представить полк в парадной форме на том основании, что всегда лучше перекланяться, чем не докланяться. И солдаты, после тридцативерстного перехода, не смыкали глаз, всю ночь чинились, чистились; адъютанты и ротные рассчитывали, отчисляли; и к утру полк, вместо растянутой беспорядочной толпы, какою он был накануне на последнем переходе, представлял стройную массу 2 000 людей, из которых каждый знал свое место, свое дело и из которых на каждом каждая пуговка и ремешок были на своем месте и блестели чистотой. Не только наружное было исправно, но ежели бы угодно было главнокомандующему заглянуть под мундиры, то на каждом он увидел бы одинаково чистую рубаху и в каждом ранце нашел бы узаконенное число вещей, «шильце и мыльце», как говорят солдаты. Было только одно обстоятельство, насчет которого никто не мог быть спокоен. Это была обувь. Больше чем у половины людей сапоги были разбиты. Но недостаток этот происходил не от вины полкового командира, так как, несмотря на неоднократные требования, ему не был отпущен товар от австрийского ведомства, а полк прошел тысячу верст.
В этой части персональных интерпретаций мы поговорим о гаплогруппах, которые некоторые называют «минорными» (по численности, разумеется), или «экзотическими», в общем, тех, которых относительно немного среди этнических русских, да и вообще северных народов. Это гаплогруппы J1, J2 и H. Среди этнических русских – напомню, что этническими русскими называют тех, для кого русский – родной язык в нескольких поколениях, чьи предки жили на территории исторической России, а именно её европейской части, в нескольких поколениях, и кто считают себя русскими, – так вот, среди этнических русских гаплогруппы J1 практически нет, в лучшем случае на уровне десятых долей процента, гаплогруппы J2 примерно 3%, и гаплогруппы Н тоже практически нет. Но это среди этнических русских, а в Росийской Федерации, напомним, живут далеко не только этнические русские, и они тоже присылают заявки на персональные интерпретации «подписей» в своих Y-хромосомах. А у них, особенно тех, кто живут на восточном Кавказе, эти гаплогруппы вовсе не экзотика. Например, у кубачинцев содержание гаплогруппы J1 достигает 98%, у даргинцев 69%, у лезгин 44%, у чеченцев 23%. Содержание гаплогруппы J2 у ингушей составляет 82%, у чеченцев 58% (в Дагестане) и 51% (в Ингушетии). Гаплогруппа Н мало у всех в РФ, кроме цыган, и отдельных вкраплений в отдельные этносы.
По какому-то уже устоявшемуся стереотипу многие считают, что гаплогруппы J1 и J2 – это еврейские гаплогруппы. Это, конечно, не так, как уже видно из перечня некоторых кавказских этносов выше, они вовсе не евреи. Общие предки гаплогруппы J2 у многих групп на Кавказе уходят на глубину времен 6-8 тысяч лет назад, и они пришли частью с так называемыми урукскими миграциями из Месопотамии, частью из Передней Азии, частью из Средиземноморья. Евреев как таковых тогда вообще не было, евреи и арабы как этно-социальные и религиозные образования появились только 4000-3600 лет назад. Так что расшифровка персональных ДНК-генеалогий носителей этих гаплогрупп может принести много интересного. Кстати, гаплогруппы J1 и J2 недавно найдены в ископаемых скелетных останках на хазарских и аланских территориях между Доном и Волгой, и когда соответствующие гаплотипы будут обнародованы, кто-то из носителей этих гаплогрупп может с удивлением узнать в них свой гаплотип, отдаленный всего на тысячу-полторы тысячи лет, то есть всего на пару мутаций в 17-19-маркерном гаплотипе. А если ингуши, например, опознают свои современные гаплотипы в аланских ископаемых гаплотипах на тех территориях, будет, наверное, большой национальный праздник. Ради таких моментов, пожалуй, и стоит заниматься ДНК-генеалогией. Ну кто еще и каким образом может принести национальные праздники дружественным народам, да такими малыми силами? Наука порой делает огромное дело не только в создании новой техники или лечении смертельных болезней, но и в подъеме национального духа. То, что я называю научным патриотизмом, как бы это ни было неприятно тем, для кого патриотизм – бранное слово.
Но есть и другие ситуации – когда открытие своих исторических корней приносит шок и расстройство. Все-таки стереотипы и национальные предрассудки – тоже большая сила, с которой надо справляться. Об одном таком случае я здесь тоже расскажу.
Гаплогруппа J1
Эту гаплогруппу в наших интерпретациях обнаружил пока один человек, представивший свой 67-маркерный гаплотип, вкупе с надеждой, что он происходит напрямую от библейского Авраама.
KLIN ID00015
Представлен 67-маркерный гаплотип, и сообщено, что он относится к субкладу J-ZS227:
12 23 14 10 13 15 11 16 12 13 11 30 – 17 8 9 11 11 26 14 21 25 12 14 16 17 – 11 10 20 22 15 14 20 19 31 36 12 10 – 11 8 15 16 8 11 10 8 11 9 12 21 22 18 10 12 12 15 8 12 24 21 13 12 12 14 12 12 12 11
С отнесением этого субклада есть сложности, потому что он есть только в классификации FTDNA (см. диаграмму ниже), но нет ни в классификации ISOGG, ни в геномной классификации компании YFull, причем между ISOGG и YFull идет полный разнобой в снипах. Классификация гаплогруппы J в ISOGG на интересующем нас участке дерева гаплотипов следующая:

Судя по обсуждениям в сети, снип ZS227 находится ниже L147.1 в цепочке снипов P58 > L92.1 > L147.1
> L222.2 > L65.2, но где именно в этой цепочке – пока неизвестно.
Еще цепочка снипов, определенно из этой же серии, но с другими названиями снипов, поэтому ее трудно состыковать с теми, что выше: J1a2-L136 > J1a2b2-L147.1 > Z2313/YSC0000234 > Z1884 > Y3015. Возраст снипов в этой цепочке определен компаний YFull, как соответственно 15900 > 8800 > 5300 > 5300 > 4600 лет, то есть все они старше предполагаемого возраста Аарона (см. ниже).
Наконец, согласно классификации FTDNA (см. диаграмму ниже), цепочка снипов, ведущая к ZS227, следующая:
J-M304 > J1-M267 > CTS12238 > Z2217 > L620 > PF4832 > L136 > P58 > Z1889 > Z1853 > Z1865 > Z2324 > PF4838 > PF4843 > CTS11741 > YSC0000234 > ZS241 > ZS227
Здесь только два снипа (выделено курсивом), соответствуют классификации ISOGG.
Третий снип от конца, YSC0000234, датирован компанией YFull как образовавшийся 5300 лет назад, опять же во времена, старше библейского Авраама. Двух последующих снипов, ZS241 и ZS227, в списке компании YFull нет, и вместо них есть длинная цепочка снипов, доходящая до времени 750 лет назад:
YSC0000234/Z2313 (5300 лн) > Z1884 (5300 лн) > Y3015 (4600 лн) > S21237 (4200 лн) > Y4348 (3300 лн) > Y4349 (3300 лн) > Y10887 (3200 лн) > Y5321 (3100 лн) > Y9271 (2400 лн) > Y5323 (2000 лн) > Y5320 (1850 лн) > Y8537 (1800 лн) > FGC4474 (1500 лн) > Y12364 (1500 лн) > FGC17804 (750 лн) > Y15612 (750 лн).
Где на этом пути находятся снипы ZS241 и искомый ZS227, которые согласно компании FTDNA находятся на пути YSC0000234 > ZS241 > ZS227, остается неизвестным, и какова их датировка – выше времен библейских Авраама и Аарона, между ними, или ниже Аарона – тоже пока остается неизвестным. Война номенклатур для обозначения снипов продолжается.

Систематика фрагмента гаплогруппы J1 в номенклатуре FTDNA
Интерес к этим цепочкам снипов вызван в первую очередь тем, что субклад J1-P58 ранее приписывался коэнам (коханим), прямым потомкам первосвященника Аарона, брата Моисея, выведшего евреев из Египта примерно 3600 лет назад. Сначала, в конце 1990-х, коэнов просто определяли в гаплогруппу J1 (правда, коэны, носители J2, были не согласны, и считали, и продолжают считать, что у Аарона была гаплогруппа J2). Затем, в ходе наиболее детальной работы 2000-х годов, Hammer et al (2009) выяснили, что по результатам тестирования 215 коэнов они расходятся по 21 субкладу многих гаплогрупп, из которых:
Почти половина, 46% (99 человек) относятся к гаплогруппе J1-P58*,
14% (31 человек) – гаплогруппе J2a-M410,
7% (16 человек) – гаплогруппе J2b-M12,
6% (13 человек) – гаплогруппе J2a-M318,
5.6% (12 человек) – гаплогруппе R1b-M269,
4.6% (10 человек) – гаплогруппе E1b-M123,
по 2.3% (по 5 человек) – в гаплогруппах E1b-M78 и R1a-M17,
1.9% (4 человека) – в гаплогруппе Н-М69
Так что, строго говоря, вопрос о принадлежности «линии Аарона» пока не разрешен, но исходя просто из доли субкладов, можно полагать, что наибольшая вероятность – это J1-P58*, в тогдашней классификации.
Потом было найдено, что субклад J1-P58 является не терминальным (и потому не должен маркироваться звездочкой), а у него есть нисходящие субклады, и таких показано четыре в классификации ISOGG (см. выше), причем ниже L147.1 – два потенциально коэнских субклада. Аарон мог быть только в одном субкладе, потому что они все древние, и намного старше времен Аарона (который жил примерно примерно 3600-3300 лет назад, если следовать библейским описаниям и их более поздним толкованиям). Но J1-P58 образовался примерно 14500 лет назад (рассчитано по снипам, то есть геномным данным, компанией YFull), субклад L147.1 – 8800 лет назад, по другим субкладам из данной цепочки датировок пока нет. Поэтому отнесение Аарона на основании «общественного мнения» происходило к субкладу L147.1, хотя это почти ни на чем не основано, кроме общих рассуждений, «по понятиям».
Но после развертывания геномных исследований и идентификации снипов, выяснилось, что под субкладом J1-L147.1 находится 78 субкладов (!), из них 27 субкладов возрастом от 3300 лет и выше, остальные 51 субклад моложе 3300 лет (данные компании YFull). Так что Аарон мог быть приписан к практически любому из тех 27 субкладов. В классификации ISOGG их пока нет. Новый субклад ZS227, показанный у ID00015, оказывается уже 79-м, и мы опять же не знаем, какая там датировка.
Итог этим вступительным описаниям можно подвести такой, что мы реально не знаем, какая линия относится к библейскому Аарону, но знаем, что коэны рассыпаны по самым разным гаплогруппам, при доминирующем J1-P58. Мы не знаем, какие из тех 99 коэнов субклада J1-P58 (Hammer et al, 2009) к какой из упомянутых 79 ДНК-линий относятся, потому что такие исследования, и тем более массовые, пока не проводили.
Представленный гаплотип формально не относится к «модальному гаплотипу коэнов», который в устаревшем 6-маркерном формате (DYS 19, 388, 390, 391, 392, 393) выглядит как 14-16-24-10-11-12. У ID00015 это 14-16-23-10-11-12, то есть на одну мутацию в сторону, но само понятие «модальный гаплотип коэнов» относится к предковому 6-маркерному гаплотипу, и не учитывает мутации, которые могли накопиться со времен Аарона. Этот термин, «модальный гаплотип коэнов», броский, но научной основы не имеет. На самом деле, «коэны» столь часто имеют эту шестерку только потому, что общий предок этой шестерки жил примерно 1050 лет назад, как было подробно показано в статье (Klyosov, Human Genetics, 2009), и со столь близких времен мутаций накопилось мало.
Поэтому сейчас проведем собственное расследование этого вопроса в применении к представленному гаплотипу. Для этого возьмем 738 гаплотипов гаплогруппы J1, причем в 111-маркерном формате, это всё, что доступно на сегодняшний день в Проекте FTDNA, оставим в них 67-маркерные гаплотипы, сняв временно панель из маркеров 68-111 (она нам пригодится позже), добавим несколько 67-маркерных гаплотипов, для которых известно глубокое типирование (под номерами 740-742), добавим представленный гаплотип, обозначив его не номером, а индексом Х, и построим дерево из 742 67-маркерных гаплотипов с помощью профессиональной компьютерной программы (PHYLIP). Цель этого эксперимента – выяснить, в окружении каких гаплотипов и из каких субкладов окажется представленный галотип.
Дерево этих 67-маркерных гаплотипов гаплогруппы J1 приведено ниже для общей иллюстрации, и далее приведена ветвь, в которой находится представленный гаплотип, в увеличенном виде.

Положение представленного гаплотипа обозначено на дереве индексом Х. Выбивающиеся из дерева гаплотипы в правом нижнем углу (номера 728, 729 и ближайшие) – не относятся к гаплогруппе J1, как указано в Проекте. Непонятно, зачем их там держат. Мы оставили их только потому, чтобы показать, насколько 67-маркерное дерево гаплотипов чувствительно к «чужакам».
На диаграмме выявленной ветви (см. ниже) представленный гаплотип находится в окружении серии гаплотипов под близкими друг к другу номерами, в основном от 630 до 643. Как показано ниже, гаплотипы, не попадающие в эту серию (18, 35, 36, 614, 742) являются здесь случайными, и 111-маркерное дерево их отбросит. Но в любом случае они не находятся рядом с представленным гаплотипом. Иначе говоря, даже 67-маркерное дерево не дает порой адекватного разрешения, во всяком случае, по сравнению с 111-маркерными гаплотипами, что, в общем, понятно.

Посмотрим, к каким субкладам относятся родственные представленному гаплотипы, под номерами 630-643. Большинство из них принадлежат евреям – Presser (634), Fisher (635), Cohen (636), Aron Shirek (637), Avigdor Kohn (638), Moshe Weisinger (639), Katz (640), Cohen (641), Gutfrajnd (642), Katz (643). Гаплотипы 630-633 – из Италии и (один, 632) из Польши. Субклады для них следующие:
Ближайшие к представленному гаплотипы – 631 и 634. Первый (гаплотип 631) имеет снип Z2313, который показан на диаграмме ниже.
J1a2-L136 > J1a2b2-L147.1 > Z2313 /YSC0000234 > Z1884 > Z640
Второй (гаплотип 634) – из блока из пяти гаплотипов 634-638, коллективно обозначенных в Проекте J1 как J-S12192, которого в классификации нет (ни в ISOGG, ни в YFull). 635-й гаплотип обозначен как J-P58, и 636-й как ZS227 (снип ID00015). 637 и 638 обозначены как Z2313/YSC0000234, его «возраст», как сообщалось выше – примерно 5300 лет.
Гаплотип 639 показан как BY68/Y5399 (образован 2500 лет назад). Гаплотипы 642 и 643 показаны как относящиеся к снипу ZS2379, которого в классификациях (помимо FTDNA) нет. Они также обозначены как Р58, видимо, показывая родительский снип.
Гаплотипы 640 и 641 (Cohen) показаны как имеющие снипы ZS2361 = Y13968, причем для гаплотипа 640 еще указано, что он относится к снипу ZS227. Но возраст снипа Y13968 по данным YFull – всего 750 лет. Если эти равенства (эквивалентности) снипов действительно верны, то у ID00015 возраст снипа ZS227 – 750 лет. Но, как будет показано ниже, это неверно, видимо, или у YFull с датировкой ошибка, или сообщение администраторов Проекта FTDNA, что ZS227 эквивалентен снипам ZS2361/Y13968, неверно. Скорее, это администраторы Проекта ошиблись.
Итак, наше рассмотрение приводит к тому, что классификация снипов в окружении представленного гаплотипа/Y-хромосомы чрезвычайно запутана. Многие снипы в классификацию еще не внесены, и родственные представленному гаплотипы, сидящие на той же ветви, имеют самые разнообразные снипы, которые могут пересекаться, но классификации это не отражают. Наконец, возможен вариант, по которому снип ID00015 имеет возраст всего 750 лет, если он действительно эквивалентен снипам ZS2361 и Y13968, но этот вариант практически невероятен (см. ниже). Ниже – воспроизведение соответствующего фрагмента из систематики компании YFull:

В верхней строке мы видим снип Z2313 (с возрастом 5300 лет), от которого последовательно отходят снип Y3081 (c таким же возрастом), далее снип Y5399 (с возрастом 2500 лет), и в последней строке – снип Y13968/ZS2361 с возрастом 750 лет. Этот последний снип имеет Cohen (гаплотип 641) и Katz (гаплотип 640).
Продолжим перекрестную проверку полученных данных. Построим дерево из тех же гаплотипов, но в 111-маркерном формате, и найдем на нем искомую ветвь, выявленную на дереве 67-маркерных гаплотипов. Если она сохранилась, значит, она стабильна и не зависит от перехода от 67-маркерных гаплотипов к 111-маркерным. Дерево приведено ниже, в сугубо иллюстративном виде.

Дерево из 738 гаплотипов гаплогруппы J1 в 111-маркерном формате.
Гаплотипа ID00015 на дереве нет, так как он был получен только в 67-маркерном формате.
Для сведения, базовый (предковый) гаплотип дерева следующий:
12 23 14 11 13 18 11 16 11 13 11 30 – 18 8 9 11 11 26 14 20 26 12 14 16 17 – 10 10 22 22 14 14 18 18 32 36 12 10 – 11 8 15 16 8 11 10 8 11 9 12 21 22 18 10 12 12 15 8 12 25 21 14 12 11 13 12 12 12 11
В нем выделены 20 мутаций, на которые отклоняется гаплотип ID00015 от базового гаплотипа J1. Это эквивалентно 20/0.12 = 167 → 200 условных поколений, или примерно 5000 лет между ними (0.12 – это константа скорости мутации для 67-маркерных гаплотипов, стрелка – поправка на возвратные мутации). Все 738 гаплотипов имеют 13074 мутаций от базового гаплотипа, это соответствует времени, когда жил общий предок современных носителей этого гаплотипа, которое составляет в данном случае 13074/738/0.12 = 148 → 174 условных поколений (по 25 лет каждое, под этот срок калибровались константы скорости мутаций), или примерно 4350±440 лет назад. На самом деле, это только грубое приближение, поскольку даже невооруженным глазом видно, что правая половина дерева моложе левой (высота ветвей справа меньше, чем слева; более тщательная обработка дерева дает возраст левой ветви, которая составлена в основном из гаплотипов евреев и арабов, примерно 4000 лет). Ясно, что это относительно недавнее время по сравнению со временем образования субклада J1 (примерно 31600 лет назад, определено по снипам компанией YFull), и это свидетельствует, что гаплогруппа J1 прошла около 4000 лет назад, то есть во времена жизни библейского Авраама, жесткое бутылочное горлышко популяции. Тогда же разошлись племена, которые в будущем составили евреев и арабов.

Выше приведена в увеличенном виде искомая ветвь, только в 111-маркерном варианте. Видно, что практически все гаплотипы из окружения представленного гаплотипа (под номерами 630-643) сохранились в ветви. Таким образом, это действительно максимально родственные гаплотипы из выборки FTDNA.
12 23 14 10 13 15 11 16 12 13 11 30 – 17 8 9 11 11 26 14 21 26 12 14 16 17 – 11 10 22 22 15 14 19 18 31 35 13 10 – 11 8 15 16 8 11 10 8 11 9 12 21 22 17 10 12 12 15 8 12 24 21 13 12 11 14 12 12 12 11
Отклоняется от «модального гаплотипа коэнов», и вместо приведенных выше 14-16-24-10-11-12 (в устаревшем 6-маркерном формате DYS 19, 388, 390, 391, 392, 393) выглядит как 14-16-23 -10-11-12. В целом ветвь отличается от представленного выше базового гаплотипа дерева на 8 мутаций (отмечено), то есть на 8/0.12 = 67 → 72 условных поколений, то есть примерно на 1800 лет. Все 14 гаплотипов в 111-маркерном формате имеют 172 мутации от 111-маркерного базового гаплотипа, что показывает, что общий предок этих 14 человек жил примерно 172/14/0.198 = 62 → 66 условных поколений, то есть 1650±210 лет назад, и общий предок представленного и этого базового (предкового) гаплотипа жил примерно (1800+1650)/2 = 1725 лет назад. Согласование этих расчетных дат находится в пределах погрешности, и еще раз подтверждает, что найдена именно ветвь ID00015, что ей примерно 1650±210 лет, и что общий предок данной ветви был прямым предком ID00015. Это означает также, что субкладу ZS227 никак не может быть 750 лет, согласно Проекту FTDNA. Этому субкладу должно быть не менее 1400-1800 лет, и все гаплотипы из искомой ветви имеют снипы или с одинаковым (например, тот же ZS227), или близким к нему возрастом, так, что вид 111-маркерного гаплотипа почти не изменился.
Исторический путь к представленному гаплотипу примерно следующий. Гаплогруппа J образовалась около 43 тысяч лет назад (из сводной гаплогруппы IJ), затем примерно 32 тысячи лет она разошлась на гаплогруппы J1 и J2, гаплогруппа J1a образовалась около 19 тыс. лет назад, и после серии субкладов образовалась гаплогруппа/субклад P58, примерно 14500 лет назад. Это было, видимо, в среде древних бедуинов или других ближневосточных кочевников, и в итоге этот субклад во множестве попал как в еврейскую, так и в арабскую среду. Это же произошло и с субкладом L147.1, который образовался примерно 8800 лет назад. Не подходит на роль субклада Аарона и Z2313, который возник 5300 лет назад, и только субклад S21237, возрастом 4200 лет, мог бы претендовать на субклад Авраама и 12 колен израилевых, включая Аарона и его потомков. Дело в том, что нижестоящие субклады Y4348 и Y4349 образовались только примерно 3300 лет назад, что уже для Авраама и его потомков поздно. Проблема в том, что на S21237, как и на эквивалентные субклады FGC1722, FGC1698, FGC1701, Y5585 и Y8768, пока тестировали очень мало.
Субклад ZS227 пока не датирован, но дерево гаплогрупы J1 (см. выше) показывает, что ветвь ID00015 образовалась 1650±210 лет назад. Возможно, что эта ветвь образовалсь от общего предка, который принадлежал к линии Аарона, но для выяснения такого варианта надо поставить в соответствие на дереве субкладов снипы S21237 (4200 лет назад) и ZS227, которые пока относятся к разным номенклатурам. Если второй – нижеследующий субклад по отношению к первому, то мы значительно приблизимся к решению загадки о происхождении коэнов от Аарона, и какие коэны (из многих) принадлежат этой линии. Возможно, ДНК-линия ID00015 тоже ведет от Аарона и ранее от Авраама, если принять имена, которые называют Тора и Библия.
Гаплогруппа J2
Эту гаплогруппу в наших интерпретациях обнаружил тоже пока один человек.
KLIN ID00017
Представлен 67-маркерный гаплотип и сообщено, что гаплотип относится к субкладу J2, более детальных сведений нет.
12 25 14 10 14 16 11 15 11 13 11 29 – 16 9 9 11 11 25 17 19 30 13 15 15 18 – 10 10 21 22 15 13 20 20 34 35 10 10 – 11 7 14 14 8 12 10 8 9 9 12 20 24 16 10 12 12 16 8 11 25 20 14 13 11 13 10 12 12 11
Вообще с субкладами гаплогруппы J в номенклатурах какая-то напасть, и с J1, с J2. Понятно, что это – человеческий фактор, что в переводе на простой русский язык означает, что соответствующие активисты не делают то, что им надлежит делать. Классификация ISOGG по гаплогруппе J2 давно устарела и практически не пополняется уже долгое время, геномная классификации компании YFull значительно более подробная, но использует другие названия субкладов, которые часто не соответствуют тем, которые есть в класификации ISOGG, а компания FTDNA использует свою номенклатуру и названия снипов. Поэтому для начала воспроизведем базовую номенклатуру ISOGG (для гаплогруппы J2a), и затем попытаемся дополнить ее теми субкладами и снипами, которые нам понадобятся для решения представленной задачи. В правой колонке представлены примерные датировки возникновения древнейших снипов, рассчитанные компанией YFull по геномным данным.

Интерес многих к гаплогруппе J2, а именно к тем субкладам, в которых преобладают евреи, вызван, в первую очередь, тем, что какие-то из этих субкладов могут быть отнесены к коэнам (коханим), прямым потомкам первосвященника Аарона. Ситуация с коэнами в рамках ДНК-генеалогии описана в предшествующей Интерпретации. В итоге, строго говоря, вопрос о принадлежности «линии Аарона» пока не разрешен. Как было отмечено выше, до настоящего времени отнесение Аарона на основании «общественного мнения» проводится к субкладу L147.1, хотя это почти ни на чем не основано, кроме общих рассуждений «по понятиям». Представленный гаплотип в 6-маркерном формате имеет вид 14-15-25-10-11-12, и отклоняется от «модального гаплотипа коэнов» 14-16-24-10-11-12 на две мутации, но это не имеет никакого значения, тем более в гаплогруппе J2.
Попытаемся определить место представленного гаплотипа на общем дереве 67-маркерных гаплотипов гаплогруппы J2 из обширного Проекта J2 FTDNA. Для этого возьмем все 417 111-маркерных гаплотипов гаплогруппы J2 Проекта , добавим к ним все 111-маркерные гаплотипы Проекта франкфуртских евреев гаплогруппы J2 (их в гаплогруппе J2 впятеро больше, чем в гаплогруппе J1), поскольку ID00017 этой ветвью особенно интересовался, оставим в них 67-маркерные гаплотипы, сняв временно панель из маркеров 68-111 (она нам пригодится позже), добавим представленный гаплотип, обозначив его индексом Х, и построим дерево из 434 67-маркерных гаплотипов с помощью компьютерной программы (PHYLIP). Цель этого эксперимента – выяснить, в окружении каких гаплотипов и из каких субкладов окажется представленный гаплотип. Дерево этих 67-маркерных гаплотипов гаплогруппы J2 приведено ниже для общей иллюстрации, и далее приведена ветвь, содержащая представленный гаплотип, в увеличенном виде.

Положение представленного гаплотипа обозначено на дереве индексом Х. Это – отдельная, хорошо выраженная ветвь подобных по виду гаплотипов, и, как мы покажем ниже, среди них имеется плотная группа родственных гаплотипов. Для сведения, базовый (предковый) гаплотип дерева следующий:
12 23 14 10 13 16 11 15 12 13 11 29 – 16 9 9 11 11 26 15 20 30 13 14 15 16 – 10 10 19 22 15 14 17 17 35 37 11 9 – 11 7 15 15 8 11 10 8 11 9 10 18 8 15 10 12 12 16 8 12 22 21 13 12 11 14 11 12 12 11
Здесь отмечены 50 мутаций, которые имеет представленный гаплотип от базового гаплотипа J2. Это эквивалентно 50/0.12 = 417 → 690 условных поколений, или примерно 17250 лет между ними (0.12 – это константа скорости мутации для 67-маркерных гаплотипов, стрелка – поправка на возвратные мутации). Все 434 гаплотипов имеют 14082 мутаций от базового гаплотипа, это соответствует времени, когда жил общий предок современных носителей этого гаплотипа, которое составляет в данном случае 14082/434/0.12 = 270 → 367 условных поколений (по 25 лет каждое, под этот срок калибровались константы скорости мутаций), или 9175±920 лет назад. Ясно, что это относительно недавнее время по сравнению со временем образования субклада J2 (примерно 31600 лет назад, определено по снипам компанией YFull), и это свидетельствует, что гаплогруппа J2 прошла около 9000 лет назад жесткое бутылочное горлышко популяции. Общий предок ID00017 и 434 носителей гаплогруппы J2 жил примерно (17250+9175)/2 = 13200 лет назад, то есть еще до прохождения бутылочного горлышка.
На увеличенной диаграмме искомой ветви (см. ниже) представленный гаплотип находится в окружении серии гаплотипов под близкими друг к другу номерами, от 330 до 340. Из них гаплотипы от 334 до 340 образуют свою плоскую ветвь, значит, ветвь относительно недавних гаплотипов, то есть гаплотипов от относительно недавнего общего предка.

Группа франкфуртских евреев (номера гаплотипов 60-80 и 417-447) находится совсем на другой части дерева, в верхней левой ветви, ветвь отмечена символом F. Характеристика этой ветви будет дана ниже, в сравнении с искомой ветвью. Пока достаточно сказать, что они от ветви ID00017 очень далеко.
Продолжим перекрестную проверку полученных данных. Построим дерево из тех же гаплотипов, но в 111-маркерном формате, и найдем на нем искомую ветвь. Если она сохранилась, значит, она стабильна и не зависит от перехода от 67-маркерных гаплотипов к 111-маркерным. Дерево приведено ниже, в сугубо иллюстративном виде. Как видно, общая форма дерева сохранилась, программа его немного развернула относительно центральной оси, что не имеет для нас никакого значения, нам важно, чтобы сохранились ветви.

Полученные выше расчеты для времени жизни общего предка современных носителей гаплогруппы J2 по 67-маркерным гаплотипам можно сопоставить с расчетами по 111-маркерному дереву. Все 433 гаплотипов имеют 25028 мутаций от базового 111-маркерного гаплотипа, это соответствует времени, когда жил общий предок современных носителей этого гаплотипа, которое составляет в данном случае 25028/433/0.198 = 292 → 408 условых поколений, или примерно 10200±1020 лет назад. Это в пределах погрешности совпадает с расчетами по 67-маркерным гаплотипам (9175±920 лет назад).

Выше приведена в увеличенном виде искомая ветвь, только в 111-маркерном варианте. Видно, что практически все гаплотипы из окружения представленного гаплотипа (под номерами 334-340) сохранились в ветви. Таким образом, это действительно максимально родственные представленному гаплотипы из выборки FTDNA. Все они очень похожи друг на друга.
Базовый гаплотип этой ветви в 67-маркерном формате
12 25 14 10 14 16 11 15 11 14 11 30 – 16 9 9 11 11 25 16 20 30 13 15 15 18 – 10 10 21 22 15 13 20 20 34 36 10 10 – 11 7 14 14 8 12 10 8 9 9 12 20 24 16 10 12 12 17 8 11 25 20 14 13 11 13 10 12 12 11
Отличается от представленного гаплотипа всего на 5 мутаций (отмечены); первые две – это одна мутация по правилам счета), то есть на 5/0.12 = 42 → 44 условных поколений, то есть примерно на 1100 лет. Все 7 гаплотипов в 111-маркерном формате имеют 54 мутации от 111-маркерного базового гаплотипа, что показывает, что общий предок этих 7 человек жил 54/7/0.198 = 39 → 41 условных поколений, то есть 1025±170 лет назад, и общий предок представленного и этого базового (предкового) гаплотипа жил примерно (1100+1025)/2 = 1060 лет назад. Согласование всех этих расчетных дат (1100, 1025, 1060 лет) находится в пределах погрешности, и еще раз подтверждает, что найдена именно ветвь ID00017, что ей 1025±170 лет, и что общий предок данной ветви был прямым предком ID00017.
Ветвь франкфуртских евреев (номера гаплотипов 60-90 из Проекта J2 FTDNA и 417-447 из Проекта Франкфуртских евреев) в увеличенном виде приведена на следующей диаграмме (напомним, что ID00017 этой ветвью специально интересовался).
Базовый (предковый) гаплотип ветви
12 22 14 10 14 15 11 15 12 14 11 31 – 15 9 9 11 11 26 15 20 29 13 13 15 16 – 10 10 19 22 15 12 15 17 34 36 12 9 – 10 7 15 15 8 11 10 8 11 9 0 17 17 15 10 12 12 15 8 11 22 21 13 12 11 13 11 12 12 11
Отличается от ветви ID00017 на 55 мутаций, то есть предковые гаплотипы обеих ветвей евреев расходятся на 55/0.12 = 458 → 800 условных поколений, или 20 тысяч лет. Поэтому они и находятся на разных сторонах дерева гаплотипов, см. выше. Все 45 гаплотипов франкфуртской ветви содержат 346 мутаций от базового гаплотипа в 111-маркерном формате, то есть общий предок ветви жил 346/45/0.198 = 39 → 41 условных поколений, или 1025±120 лет назад, то есть ровно столько же, сколько общий предок ветви ID00017. Таким образом, общий предок обеих ветвей жил (20000+ 1025+1025)/2 = 11025 лет назад.
Это, безусловно, очень необычный случай – когда две недавние семьи (в генеалогическом смысле) евреев имеют общего предка (в одной гаплогруппе) 11 тысяч лет назад, когда евреев и в помине не было. Иначе говоря, две популяции гаплогруппы J2, расходясь от общего предка, который жил 11 тысяч лет назад, стали полностью еврейскими примерно тысячу лет назад. Наверное, наиболее разумное объяснение, что они в конце 10-го века (плюс-минус век) стали «конвертами» в иудаизм.
Посмотрим, к каким субкладам относятся родственные представленному гаплотипы, под номерами 334-340. Большинство из них принадлежат евреям – Столбин, Украина (334), Mendel Siedlecki, Польша (335), Jacob Frolich, Прага (336), Juda Levin, Польша (337), Harris Sklar, ашкенази, Латвия (338), Movsha Firksler, Латвия (339), Abraham Halle, Германия (340).
Субклады у всех них в Проекте FTDNA обозначены как PF4610 > Z6049 > Z6048. К сожалению, только PF4610 удается идентифицировать, это J2a1-L26, который образовался примерно 18200 лет назад. Остальные в ISOGG и YFull не числятся. Нам же желательно найти субклад, с датировкой несколько ранее 1200 лет назад. Возможно, некоторую подсказку даст диаграмма с сайта компании YFull:

Видно, что в данной ветви, давностью 7600 лет назад, которая затем расходится на несколько подветвей 2600 и 1800 лет назад (например, на две подветви, Y6957 и Y7291, примерно 2600 лет назад), все почти исключительно «русские», то есть живущие в России. Возможно, ID00017 и имеет один из этих снипов.
Исторический путь к представленному гаплотипу примерно следующий. Начальный этап эволюции гаплогруппы J и ее расхождение на J1 и J2 описано в предыдущей Интерпретации. В итоге гаплогруппа J2 (как и J1) во множестве попала как в еврейскую, так и в арабскую среду, а также во множестве разошлась по всему Средиземноморью. В ходе Рассеяния евреи расселились по всей Европе, с очагами (часто временными) в Испании, Италии, Германии. Переходы в иудаизм в Европе были, видимо, крайне редкими. Возможно, общий предок ветви ID00017, как и ветви франкфуртских евреев, был «конвертом», и это было примерно в 10-м веке, до больших неприятностей в еврейских сообществах в Англии, Испании, Франции и вообще в Центральной Европе, особенно, во времена чумы в Европе. ID00017 – потомок одной из этих линий.
Надо отметить, что к кавказским гаплотипам гаплогруппы J2 представленный гаплотип имеет весьма малое отношение, разве что относится к той же гаплогруппе. На краях искомой (см. выше) ветви сидят гаплотипы (331, 332, 333) кавказцев Тавасилова (Северная Осетия), Ахвледиани (Грузия) и Будаева (Осетия, дигорец), но они имеют общего предка 3725±560 лет назад, и представленный гаплотип от них очень удален.
Гаплогруппа H
Эту гаплогруппу в наших интерпретациях пока показали два человека, один с 111-маркерным гаплотипом, другой с 67-маркерным.
KLIN ID00023
Представлен 111-маркерный гаплотип, сообщено, что его субклады Н-М69 и М370, место жизни предков по мужской линии – Нахичеваньская АР, Азербайджан.
12 23 15 10 14 15 11 12 10 14 11 30 – 16 9 9 11 11 24 14 18 26 12 12 13 15 – 10 11 19 21 16 14 16 18 32 37 13 11 – 11 8 16 16 8 10 10 8 12 9 12 23 23 20 9 12 12 16 8 13 22 21 14 12 11 12 9 12 14 11 – 36 16 8 15 12 21 29 19 12 11 12 12 11 9 11 12 10 11 13 31 11 13 22 16 10 11 19 15 22 11 23 14 13 15 26 12 21 20 9 15 19 9 12 11
Гаплотип относится к древнему субкладу Н1, который образовался примерно 45 тысяч лет назад. От него в дальнейшем отошло множество нижестоящих субкладов, как показано на следующей диаграмме. Помимо них имеются параллельные субклады Н2 и Н3, но они нас в данном случае не интересуют.
Было бы важно узнать, к какому из указанных субкладов относится представленный гаплотип, но поверхностное тестирование ID00023 на снипы, которое дало только Н1, такой возможности нам пока не дает. Дело в том, что все нижестоящие субклады тоже дают при типировании (то есть при идентификации снипов) Н1-М69, и все они показывают родительский субклад Н-L901, поэтому мы не знаем, представленный субклад именно М69, или любой другой из нижестоящих. Поэтому построим дерево 67-маркерных гаплотипов гаплогруппы Н, и посмотрим, на какую ветвь уйдет представленный гаплотип. Мы для начала взяли 67-маркерный формат, потому что 111-маркерных гаплотипов гаплогруппы Н мало, и дерево бедное. Но мы к нему еще вернемся. Дерево 67-маркерных гаплотипов приведено ниже, на нем гаплотип ID00023 обозначен буквой Х. Все остальные гаплотипы взяты из Проекта FTDNA гаплогруппы Н .

Видно, что представленный гаплотип входит в самую древнюю группу. Чем выше/дальше отходят гаплотипы (в виде линий на дереве), тем они более мутированы, значит – происходят от более древних общих предков. Чем ветвь компактнее, плотнее, тем более недавно жил общий предок. Самая древняя ветвь дерева расположена справа. Это – субклад Н1-М69 с вкраплениями H1a-M52. Вкрапления означают, что некоторые (или многие) 67-маркерные гаплотипы субкладов М69 и М52 настолько похожи, что программа их разделить не может, и группирует в одну ветвь. Посмотрим на 111-маркерное дерево:

Мы видим, что большинство более коротких гаплотипов древней ветви – под номерами 6, 8, 11 и 12 – не вошли в 111-маркерное дерево, но ID00023 продолжает оставаться в древней ветви, и продолжает образовывать пару с гаплотипом 13. Значит, это стабильные признаки. Но в любом случае, задачу надо решать только прямым тестированием на глубокие снипы, что будет возможно в московской Лаборатории ДНК-генеалогии, когда она откроется.
Гаплотипов вышестоящего, «корневого» субклада Н-L901 в базе данных нет (он образовался примерно 48 тысяч лет назад, и его носители могли за это время не выжить, или все получили нижестоящие снипы). Субклад М69, который выявлен у ID00023, представляют в основном современные жители Ближнего Востока и Аравии – Саудовской Аравии (треть от всех), Ирака, Кувейта, а также один пакистанец.
Если более конкретно по древней 67-маркерной ветви (справа на дереве гаплотипов), то гаплотип под номером 6 – из Ирака, под номером 8 – из Кувейта, гаплотип 10 страну в базе данных не указал, гаплотип 11 – Саудовская Аравия, 12 – Пакистан, 13 – Саудовская Аравия. Последний гаплотип, из Саудовской Аравии, ближе всех к гаплотипу ID00023 (и на 67-, и на 111-маркерном дереве) и образует с ним пару, но он – из субклада H1a-M52. Таким образом, остается неясным, ID00023 относится к субкладу М69 или М52. Как указано выше, московская Лаборатория ДНК-генеалогии будет определять эти снипы, как и практически все нисходящие, приведенные на диаграмме выше по тексту.
Посмотрим, насколько далеко от ID00023 находится гаплотип 13, образующий с ним пару. Между ними – 38 мутаций на 67 маркерах, что помещает общего предка этих двух гаплотипов на 5725±1090 лет назад. На 111 маркерах между ними ровно вдвое больше – 76 мутаций, то есть растояние между двумя гаплотипами составляет 76/0.198 = 384 → 606 условных поколений, или 15150 лет, и общий предок жил примерно 7575±1150 лет назад, что в пределах погрешности с датировкой по 67-маркерным гаплотипам. Погрешности велики, потому что в расчетах в данном случае используются всего два гаплотипа, но результаты более надежные для 111-маркерных гаплотипов. В любом случае, родства в обычном понимании этого слова, конечно, нет. Возможно, прямой предок ДНК-линии ID00023 прибыл на Кавказ со стороны Аравийского полуострова или Месопотамии во времена урукских миграций на Кавказ, около 7 тысяч лет назад.
Общий предок ветви ID00023 из семи гаплотипов (в 67-маркерном формате), в основном, из арабских стран Ближнего Востока, жил 9300±1100 лет назад. Обычно принято считать, что гаплогруппа Н зародилась в Индии, но эти данные позволяют выдвинуть альтернативную гипотезу о ближневосточном или аравийском ее происхождении, с последущими миграциями в Индостан. Если добавить к 12 гаплотипам H1-M69 в базе данных все три доступных гаплотипа нижестоящего субклада М52 (два из Индии и один из Саудовской Аравии), то датировка времени жизни общего предка будет немного моложе (формально – всего на пять поколений ближе к нам), но в пределах погрешности расчетов – 9175±1010 лет назад. Базовый гаплотип ветви из семи гаплотипов следующий:
13 22 15 10 14 16 11 12 11 13 11 30 – 16 9 9 11 11 24 14 19 29 14 14 15 16 – 10 10 19 21 14 14 16 18 35 39 11 10 – 11 8 16 16 8 10 10 8 12 10 10 22 23 16 10 12 12 16 8 12 24 20 15 12 11 13 10 11 14 11
Он отличается от гаплотипа ID00023 на 42 мутации, что дает время, когда жил общий предок ветви и представленного гаплотипа 6600±1200 лет назад, что в пределах погрешности расчетов такое же, как и время жизни общего предка с соседним гаплотипом под номером 13 как по 67-, так и по 111-маркерным гаплотипам. А сама ветвь – давностью примерно 9 тысяч лет. Вот из каких глубин времени поднимается эстафета поколений от прямых предков ID00023 к настоящему времени.
KLIN ID00008
Представлен 67-маркерный гаплотип, гаплогруппа не сообщена.
12 22 15 10 15 18 11 12 11 14 11 30 – 19 9 9 10 12 24 14 19 30 12 12 14 16 – 11 11 19 21 15 11 19 16 38 38 12 9 – 12 8 16 16 8 12 10 8 11 9 0 19 21 18 10 12 12 16 8 14 22 20 16 12 11 13 9 12 14 11
Признаться, расшифровка отнесения этого гаплотипа представила некоторые трудности, поскольку гаплогруппа Н в Европейской части является довольно редкой, и распространена в основном среди цыган. Это – гаплогруппа/субклад Н1а1, снип М82.
H L901
. H1 M69
. . H1a M52
. . . H1a1 M82
Этот субклад уходит глубоко вниз по дереву субкладов (то есть к более недавним временам), как будет показано ниже, в конце данного материала. Перед вами дерево из 54 гаплотипов гаплогруппы H в 67-маркерном формате. Представленный гаплотип ID00008 обозначен индексом Х.

Видно, что гаплотип ID00008 входит в плотную группу (слева), что означает, что общий предок этой группы жил относительно недавно. Насколько недавно – мы рассмотрим чуть позже, а пока взглянем на дерево более пристально. Чем выше отходят гаплотипы, тем они более мутированы, значит – происходят от более древних общих предков. Чем ветвь компактнее, плотнее, тем более недавно жил общий предок.
Слева (в нижней части) и внизу на дереве – гаплотипы из Индии, Пакистана, Турции, которые имеют субклад М82. Это – субклад ID00008, но более старая ветвь и с другой географией. На 13 гаплотипов этой группы приходится 214 мутаций от базового (предкового) гаплотипа
12 22 15 10 15 17 11 12 11 14 11 30 – 17 9 9 10 12 23 14 19 32 12 12 14 15 – 10 11 19 21 15 11 17 17 34 37 12 9 – 11 8 16 16 8 10 10 8 11 9 13 21 21 19 10 12 12 15 8 13 23 20 16 12 11 12 9 12 14 11
Что дает «возраст» общего предка этой группы 214/13/0.12 = 137 → 159 условных поколений, то есть 3975±480 лет. Здесь выделены 27 мутаций от представленного гаплотипа, что дает расхождение во времени между ним и предковым указанной группы того же субклада, приблизительно равное 27/0.12 = 225 → 289 условных поколений (стрелка – поправка на возвратные мутации), или 7225 лет. Иначе говоря, общий предок представленного гаплотипа и этой группы гаплотипов того же субклада жил половину этого срока назад, или примерно 3610 лет назад. Это, действительно, в пределах погрешности расчетов с величиной 3975±480 лет, приведенной выше. Кстати, если выделить из этой группы только индийские гаплотипы, то их общий предок жил 4750±610 лет назад. Мы можем заключить, что индийские гаплотипы могут быть старше турецких, хотя все это в пределах погрешности расчетов.
Мы уже вплотную приблизились к относительно недавним ветвям того же субклада Н1а1-М82. Принято считать, и к этому есть основания, что субклад М82 был принесен в Европу цыганами, или рома, как их сейчас все чаще называют на Западе, примерно 1000-1200 лет назад, в 9-11 вв. нашей эры, и их потомки, многие из которых сейчас уже не цыгане, во всяком случае, по жизненному укладу, имеют общих предков (по регионам), которые жили 900-600 лет назад. Примеры этого мы видим и на дереве гаплотипов, приведенном выше, в виде трех отдельных групп гаплотипов.
В одну группу (субклад М82) входит представленный гаплотип ID00008 (номера гаплотипов 30, 34-36, 38, 39 и Х). Эти гаплотипы предоставили их носители с предками из Украины (гаплотип 30), Италии (35), Англии (36, 37 – это дубль, который не засчитывается), Болгарии (39) и Молдовы (Х), остальные данные не предоставили. Все семь гаплотипов имеют 39 мутаций от базового гаплотипа
12 22 15 10 15 17 11 12 11 14 11 30 – 18 9 9 10 12 24 14 19 31 12 12 14 15 – 11 11 19 21 15 11 18 16 35 38 12 9 – 11 8 16 16 8 10 10 8 11 9 0 20 21 18 10 12 12 16 8 14 23 20 16 12 11 12 9 12 14 11
(12 мутаций с представленным гаплотипом выделены), что дает 39/7/0.12 = 46 → 48 условных поколений, или 1200±230 лет до общего предка ветви ID00008. Проверим дополнительно принадлежность гаплотипа ID00008 к этой ветви. 12 мутаций, которые отделяют от нее представленный гаплотип, дают 12/0.12 = 100 → 111 условных поколений, или примерно 2775 лет. Таким образом, общий предок представленного гаплотипа и указанной ветви жил примерно 2775/2 = 1390 лет назад, что попадает в коридор погрешности со временем жизни общего предка ветви – 1200±230 лет назад. Таким образом, гаплотип ID00008 действительно попадает на указанную ветвь.
Итак, идентифицирована ветвь гаплогруппы H-M82, которая именуется цыганской в Проекте FTDNA, и показано, что ее общий предок жил 1200±230 лет назад. Эти времена действительно соответствуют началу появления цыган в Европе, европейские цыгане в своим большинстве имеют гаплогруппу Н, и представленный гаплотип попадает в эту ветвь.
Соседняя с описываемой ветвью группа (гаплотипы 41-44 и 52-52, все относятся к субкладу М82) состоит полностью из английских гаплотипов, и имеет возраст несколько моложе – там на шесть гаплотипов приходится 27 мутаций, что дает 27/6/0.12 = 38 → 40 условных поколений, или 1000±220 лет до общего предка.
Наконец, ветвь из девяти гаплотипов (все тоже субклада М82, номера 31-33, 45-50) в верхней правой стороне дерева состоит из шотландских гаплотипов, за исключением гаплотипа 47, из Мексики. Он несколько выходит за пределы ветви, или, скорее, лежит в ее основании. Все девять гаплотипов имеют 35 мутаций, что дает 35/9/0.12 = 32 → 33 условных поколений, или 825±160 лет до общего предка. Если удалить мексиканский гаплотип, то получим 675±150 лет до общего предка для шотландцев субклада М82.
Таким образом, картина достаточно ясна и наглядна. Корни ДНК-линии ID00008, скорее всего, в цыганах, которые несколько более тысячелетия назад пришли из Индии. Если в его семейных преданиях о цыганах-предках ничего не известно, то этот субклад мог быть приобретен кружным путем, или просто тайным. Обращение к ID00008 – поговорите со стариками, вашими родствениками.
Как можно прояснить тайны родословной ID00008, если цыганское происхождение действительно оказалось сюрпризом, что бывает? Как мы заключили, представленный субклад, скорее всего, Н1а-М82. «Скорее всего» здесь стоит потому, что наиболее надежным будет прямое тестирование на глубокие снипы. Последовательность их от M82 и выше (по времени ближе к нам) следующая:

Большинство из этих снипов будут определяться в московской Лаборатории ДНК-генеалогии, или их можно определить, например, в FTDNA. Проблема в том, что в FTDNA предстоит играть в дорогостоящую лотерею, поскольку все снипы там определяются раздельно, каждый стоит 39 долларов, и «играющему» там никто не подскажет, какой очередной снип выбрать. Если «игроку» сильно не повезет, и все снипы будут тестироваться по одному, по очереди или вразбивку, то тестирование всех 29 снипов будет стоить 1131 доллар. В московской Лаборатории ДНК-генеалогии – максимум 300 долларов, но, возможно, и меньше, и определят сразу все снипы в один заход.
Послесловие
Как и предполагалось, цыганское происхождение оказалось для ID00008 полным сюрпризом, причем поначалу крайне неприятным. Он написал мне большое письмо, в котором сожалел, что вообще об этом узнал, и выразил сомнение в целесобразности ДНК-тестирования вообще, типа «меньше знаешь – лучше спишь». В последующей переписке он сообщил, что об отце он вообще хороших воспоминаний не имеет, потому что тот бросил их с матерью, когда мой респондент был маленьким, но он помнит, что отец был очень смуглым. Я со своей стороны писал о пагубности стереотипов, в том числе, и о цыганах, и посоветовал принять его происхождение по мужской линии как данность, и найти в этом позитивные стороны, и даже определенный шарм. Посоветовал посетить цыганский табор, или несколько, благо мой респондент живет в Молдавии, поговорить с его обитателями, послушать их песни и посмотреть танцы и пляски, и найти в них самобытность и прелесть. В общем, войти в равновесие с данностью своего происхождения, а не отталкивать его с неприятными чувствами. Судя по последующим ответам моего респондента, так оно и произошло.
Передвижениями народов (J1* ) и позже миграциями семито-говорящего населения Ближнего Востока в Испанию , Пакистан и другие регионы.
Субклады
Известные субклады J1 (M267) и соответствующие им SNP мутации по ISOGG-2010:
- J1 M267
- J1* -
- J1a M62
- J1b M365
- J1c L136
- J1c* -
- J1c1 M390
- J1c2 P56
- J1c3 P58
- J1c3* -
- J1c3a M367, M368
- J1c3b M369
- J1c3c L92, L93
- J1c3d L147
- J1c3d* -
- J1c3d1 L222
- J1c3d1* -
- J1c3d1a L65.2/S159.2
- J1c3d1* -
- J1 M267
Даже по результатам раннего коммерческого тестирования сложилось впечатление, что большинство современных носителей данной гаплогруппы относятся именно к подгруппе J1c3 - ветвь P58 , что уже ныне подтвердилось более широким тестированием и является очевидным фактом.
Распространение
Ближний Восток
Самая многочисленная подгруппа J1c3 (P58) распространёна в значительной степени среди евреев и населения Аравийского полуострова . Так же гаплогруппа в целом широко распространена в Леванте и среди семитоязычного населения Северной Африки , например ассирийцев, палестинцев-христиан, друзов. Для евреев характерны субклады J1c3* и J1c3d* (коэны). Например корневой субклад J1* (M267), будучи характерным для ассирийцев , народов Дагестана , Чечни , европейцев, у евреев и арабов практически не наблюдается.
Наибольшая концентрация данной гаплогруппы, субклад J1c3d* , наблюдается в Йемене (76 %) и Саудовской Аравии (64 %) , среди палестинцев (38 %) , в Сирии (30 %) и Ливане (25 %) .
В Восточной Анатолии значительно представлен субклад Z1842.
Частота гаплогруппы J1 заметно падает на границах арабских стран и Дагестана с другими странами, такими как Иран (10,5 %) и Турция (9 %) .
Северная Африка
Частота J1 в Северной Африке: Алжир (22 %), Египет (21 %), Тунис (17 %), Марокко (7 %).
Кавказ
В регионе Кавказа наиболее распространён макросубклад Z1842. Высокая концентрация этой гаплогруппы, особенно указанного Z1842, наблюдается среди даргинцев (84 %), аварцев (67 %), лезгин (44 %), чеченцев (35 %), кумыков (21 %).
Европа
В целом частота J1 в Европе не высока. Однако относительно высокие частоты были зафиксированы в центральных Адриатических районах Италии Горгано (англ. ) (17,2 %), Пескара (15 %), Паола (11,1 %), южносицилийской Рагузе (10,7 %) , на Мальте (8 %), Кипре (6 %) . Европейские субклады принадлежат к древнему неолитическому населению Европы, и значительно отличаются от арабских и еврейских субкладов по временным дистанциям до общего предка В Наиболее представленным в Европе, в том числе и Восточной, является субклад L1128 и его подветвь L1189, хотя на Европейском театре ДНК исследований периодически наблюдаются и макросубклад Z1842, который сам по себе преимущественно распространен в Восточной Анатолии и на Кавказе. Обнаружена исключительно восточно-европейская подветвь L1189, характерная для славянских народов.
Известные представители гаплогруппы J1
- Армянские князья Мелик-Шахназаряны ; макросубкалад Z1842.
- Дворянский шотландский род Грэмов - маркизы и герцоги Монтроуз; макросубклад P58.
Напишите отзыв о статье "Гаплогруппа J1 (Y-ДНК)"
Примечания
Ссылки
- Y-DNA_J project of FTDNA
| Эволюционное древо гаплогрупп Y-хромосомы человека | ||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
|
||||||||||||||||||||||||||||||||
| Y-хромосомный Адам | ||||||||||||||||||||||||||||||||
| A00 | A0-T | |||||||||||||||||||||||||||||||
| A1a | A1b | |||||||||||||||||||||||||||||||
| A1b1 | ||||||||||||||||||||||||||||||||
| GHIJK | ||||||||||||||||||||||||||||||||
| HIJK | ||||||||||||||||||||||||||||||||
| IJK | ||||||||||||||||||||||||||||||||
|
Но он ничего не мог ей ответить, ни, тем более, что-то предложить. Он просто был мёртв, и о том, что бывает «после», к сожалению, ничего не знал, так же, как и все остальные люди, жившие в то «тёмное» время, когда всем и каждому тяжелейшим «молотом лжи» буквально вбивалось в голову, что «после» уже ничего больше нет и, что человеческая жизнь кончается в этот скорбный и страшный момент физической смерти... – Папа, мама, и куда мы теперь пойдём? – жизнерадостно спросила девчушка. Казалось, теперь, когда все были в сборе, она была опять полностью счастлива и готова была продолжать свою жизнь даже в таком незнакомом для неё существовании. – Ой, мамочка, а моя ручка прошла через скамейку!!! А как же теперь мне сесть?.. – удивилась малышка. Но не успела мама ответить, как вдруг прямо над ними воздух засверкал всеми цветами радуги и начал сгущаться, превращаясь в изумительной красоты голубой канал, очень похожий на тот, который я видела во время моего неудачного «купания» в нашей реке. Канал сверкал и переливался тысячами звёздочек и всё плотнее и плотнее окутывал остолбеневшую семью. – Я не знаю кто ты, девочка, но ты что-то знаешь об этом – неожиданно обратилась ко мне мать. – Скажи, мы должны туда идти? – Боюсь, что да, – как можно спокойнее ответила я. – Это ваш новый мир, в котором вы будете жить. И он очень красивый. Он понравится вам. Мне было чуточку грустно, что они уходят так скоро, но я понимала, что так будет лучше, и, что они не успеют даже по настоящему пожалеть о потерянном, так как им сразу же придётся принимать свой новый мир и свою новую жизнь... – Ой, мамочка, мама, как красиво!!! Почти, как Новый Год!.. Видас, Видас, правда красиво?! – счастливо лепетала малышка. – Ну, пойдём-те же, пойдёмте, чего же вы ждёте! Мама грустно мне улыбнулась и ласково сказала: – Прощай, девочка. Кто бы ты ни была – счастья тебе в этом мире... И, обняв своих малышей, повернулась к светящемуся каналу. Все они, кроме маленькой Кати, были очень грустными и явно сильно волновались. Им приходилось оставлять всё, что было так привычно и так хорошо знакомо, и «идти» неизвестно куда. И, к сожалению, никакого выбора у них в данной ситуации не было... Вдруг в середине светящегося канала уплотнилась светящаяся женская фигура и начала плавно приближаться к сбившемуся «в кучку» ошарашенному семейству. – Алиса?.. – неуверенно произнесла мать, пристально всматриваясь в новую гостью. Сущность улыбаясь протянула руки к женщине, как бы приглашая в свои объятия. – Алиса, это правда ты?!.. – Вот мы и встретились, родная, – произнесло светящее существо. – Неужели вы все?.. Ох, как жаль!.. Рано им пока... Как жаль... – Мамочка, мама, кто это? – шёпотом спросила ошарашенная ма-лышка. – Какая она красивая!.. Кто это, мама? – Это твоя тётя, милая, – ласково ответила мать. – Тётя?! Ой как хорошо – новая тётя!!! А она кто? – не унималась любопытная девчушка. – Она моя сестра, Алиса. Ты её никогда не видела. Она ушла в этот «другой» мир когда тебя ещё не было. – Ну, тогда это было очень давно, – уверенно констатировала «неоспоримый факт» маленькая Катя... Светящаяся «тётя» грустно улыбалась, наблюдая свою жизнерадостную и ничего плохого в этой новой жизненной ситуации не подозревавшую маленькую племянницу. А та себе весело подпрыгивала на одной ножке, пробуя своё необычное «новое тело» и, оставшись им совершенно довольной, вопросительно уставилась на взрослых, ожидая, когда же они наконец-то пойдут в тот необыкновенный светящийся их «новый мир»... Она казалась опять совершенно счастливой, так как вся её семья была здесь, что означало – у них «всё прекрасно» и не надо ни о чём больше волноваться... Её крошечный детский мирок был опять привычно защищён любимыми ею людьми и она больше не должна была думать о том, что же с ними такое сегодня случилось и просто ждала, что там будет дальше. Алиса очень внимательно на меня посмотрела и ласково произнесла: – А тебе ещё рано, девочка, у тебя ещё долгий путь впереди... Светящийся голубой канал всё ещё сверкал и переливался, но мне вдруг показалось, что свечение стало слабее, и как бы отвечая на мою мысль, «тётя» произнесла: – Нам уже пора, родные мои. Этот мир вам уже больше не нужен... Она приняла их всех в свои объятия (чему я на мгновение удивилась, так как она как бы вдруг стала больше) и светящийся канал исчез вместе с милой девочкой Катей и всей её чудесной семьёй... Стало пусто и грустно, как будто я опять потеряла кого-то близкого, как это случалось почти всегда после новой встречи с «уходящими»... – Девочка, с тобой всё в порядке? – услышала я чей-то встревоженный голос. Кто-то меня тормошил, пробуя «вернуть» в нормальное состояние, так как я видимо опять слишком глубоко «вошла» в тот другой, далёкий для остальных мир и напугала какого-то доброго человека своим «заморожено-ненормальным» спокойствием. Вечер был таким же чудесным и тёплым, и вокруг всё оставалось точно так же, как было всего лишь какой-то час назад... только мне уже не хотелось больше гулять. | ||||||||||||||||||||||||||||||||
The first J1 men lived in the Late Upper Paleolithic, shortly before the end of the last Ice Age. The oldest identified J1 sample to date comes from Satsurblia cave (c. 13200 BCE) in Georgia (Jones et al. (2015)), placing the origins of haplogroup J1 in all likelihood in the region around the Caucasus, Zagros, Taurus and eastern Anatolia during the Upper Paleolithic.
Like many other successful lineages from the Middle East, J1 is thought to have undergone a major population expansion during the Neolithic period. Chiaroni et al. (2010) found that the greatest genetic diversity of J1 haplotypes was found in eastern Anatolia, near Lake Van in central Kurdistan. Eastern Anatolia and the Zagros mountains are the region where goats and sheep were first domesticated, some 11,000 years ago. Chiaroni et al. estimated that J1-P58 started expanding 9,000 to 10,000 years ago as pastoralists from the Fertile Crescent. Although they did not analyze the other branches, it is likely that all surviving J1a1b (L136) lineages share the same origin as goat and sheep herders from the Taurus and Zagros mountains.
The mountainous terrain of the Caucasus, Anatolia and modern Iran, which wasn"t suitable for early cereal farming, was an ideal ground for goat and sheep herding and catalyzed the propagation of J1 pastoralists. Having colonised most of Anatolia, J1 herders would have settled the mountainous regions of Europe, including the southern Balkans, the Carpathians, central and southern Italy (Apennines, Sicily, Sardinia), southern France (especially Auvergne), and most of the Iberian peninsula. Hotspots of J1 in northern Spain (Cantabria, Asturias) appear to be essentially lineages descended from these Southwest Asian Neolithic herders.
Most J1 Europeans belong to the J1-Z1828 branch , which is also found in Anatolia and the Caucasus, but not in Arabic countries. The Z1842 subclade of Z1828 is the most common variety of J1 in Armenia and Georgia. There are also two other minor European branches: J1-Z2223 , which has been found in Anatolia and Western Europe, and J1-M365.1 , also found a bit everywhere across Western Europe. Their very upstream position in the phylogenetic tree and their scarcity in the Middle East suggests that these were among the earliest J1 lineages to leave the Middle East, perhaps as Late Paleolithic or Mesolithic hunter-gatherers that wandered outside Anatolia and, pushed by successive waves of migrations from the Neolithic to the Bronze Age, ended up in Western Europe.
Within the Middle East, SNP analysis shows that the J1-L136 branch migrated south from eastern Anatolia and split in four directions: Anatolia/Europe (PF7263), the Levant, the southern Zagros (and southern Mesopotamia ?), and the mountainous south-western corner of the Arabian peninsula (mostly in Yemen), bypassing the Arabian Desert. That latter group, consisting essentially of J1-P56 lineages, crossed the Red Sea to settle Sudan, Eritrea, Djibouti and northern Somalia. The climate would have been considerably less arid than today during the Neolithic period, allowing for a relatively easy transmigration across the Middle East with herds of goats.
Neolithic J1 goat herders were almost certainly not homogenous tribes consisting exclusively of J1 lineages, but in all likelihood a blend of J1 and lineages. So much is evident from the presence of both J1 and T1 in north-east Africa, Yemen, Saudi Arabia, but also in the Fertile Crescent, the Caucasus and the mountainous parts of southern Europe. Maternal lineages also correlate. Wherever J1 and T1 are found in high frequency, mtDNA haplogroups , N1 and are also present, as well as J, K and T to a lower extent (=>see ). It is unclear whether goats were domesticated by a tribe that already comprised both J1 and T1 lineages, or if the merger between the two groups happened during the Neolithic expansion, when two separate tribes would have bumped into each others, intermixed, and thereafter propagated together.
Bronze Age expansion of J1-P58
Kitchen et al. (2009) estimated through a Bayesian phylogenetic analysis that Semitic languages originated in the Levant around 3,750 BCE, during the Early Bronze Age. It evolved into three groups: East Semitic (an extinct branch that comprised Akkadian), Central Semitic (which gave rise to Aramaic, Ugaritic, Phoenician, Hebrew and Arabic), and South Semitic (South Arabian and Ethiopian).
J1-P58, the Central Semitic branch of J1, appears to have expanded from the southern Levant (Israel, Palestine, Jordan) across the Arabian peninsula during the Bronze Age, from approximately 3,500 to 2,500 BCE. Camels were domesticated in Somalia and southern Arabia c. 3,000 BCE, but did not become widely used in the southern Levant before approximately 1,100 BCE. Camels played an important role in the further diffusion of J1-P58 lineages, notably with the Bedouins in the desertic parts of the Middle East and North Africa. Bedouins now make up a substantial percentage of the population of Sudan (33%), Libya (15%), the United Arab Emirates (8%) and Saudi Arabia (5%).
The two most common Jewish subclades of J1 downstream of P58 are Z18297 and ZS227 . The latter includes the Cohanim haplotype. Most of the other branches under P58 could be described as Semitic, although only FGC12 seems to be genuinely linked to the medieval Arabic expansion from Saudi Arabia.
Based on very limited data, the main Lebanese subclades of J1 appear to be J1-Z640 and J1-YSC76 . Both subclades have also been found in Sicily, Andalusia and Portugal, which suggests that they were already found among the Phoenicians . However, since the Arabs conquered the same regions as those colonised by the Phoenicians, it is too early to reach such a conclusion. Subclades found in Sardinia are very useful as practically all J1-P58 on the island were supposedly brought by the Phoenicians. They include Z18297 (could also be Jewish), Z2324>YP4763, YSC76>FGC15940>ZS1690 and YSC76>FGC8223>FGC8216>FGC8196.
Geographic distribution
Distribution of haplogroup J1 in Europe, the Middle East & North Africa
Frequencies of haplogroup J1 in Europe and West Asia tend to vary considerably from one regional community to the next. The highest local percentages in Europe are found in Greece, Italy, France, Spain and Portugal and hardly ever exceed 5% of the population. However Italy, France and Spain also have areas where J1 appears completely absent. Even in northern Europe, where the nation-wide frequencies are below 0.5%, very localised pockets of J1 have been observed in Scotland, England, Belgium, Germany and Poland. Larger sample sizes are needed to get a clearer picture of the distribution of J1 in Europe.
Surprisingly, even in the Caucasus and in Anatolia, the region where this haplogroup is thought to have originated, there are wide discrepancies between regions. For example, the Kubachi and Dargins from Dagestan in the Northeast Caucasus have over 80% of J1 lineages, while in their Ingush neighbours, 200 km to the north, it barely reaches 3%. East Anatolia around Lake Van sees over 30% of J1, whereas south-west Anatolia has only 2%. Even within Kurdistan frequencies vary greatly. The small sample sizes for each region is surely to blame.
In Arabic countries, J1 climaxes among the Marsh Arabs of South Iraq (81%), the Sudanese Arabs (73%), the Yemeni (72%), the Bedouins (63%), the Qatari (58%), the Saudi (40%), the Omani (38%) and the Palestinian Arabs (38%). High percentages are also observed in the United Arab Emirates (35%), coastal Algeria (35%), Jordan (31%), Syria (30%), Tunisia (30%), Egypt (21%) and Lebanon (20%). Most of the Arabic J1 belongs to the J1-P858 variety.
Phylogeny of J1
If you are new to genetic genealogy, please check our Introduction to phylogenetics to understand how to read a phylogenetic tree.
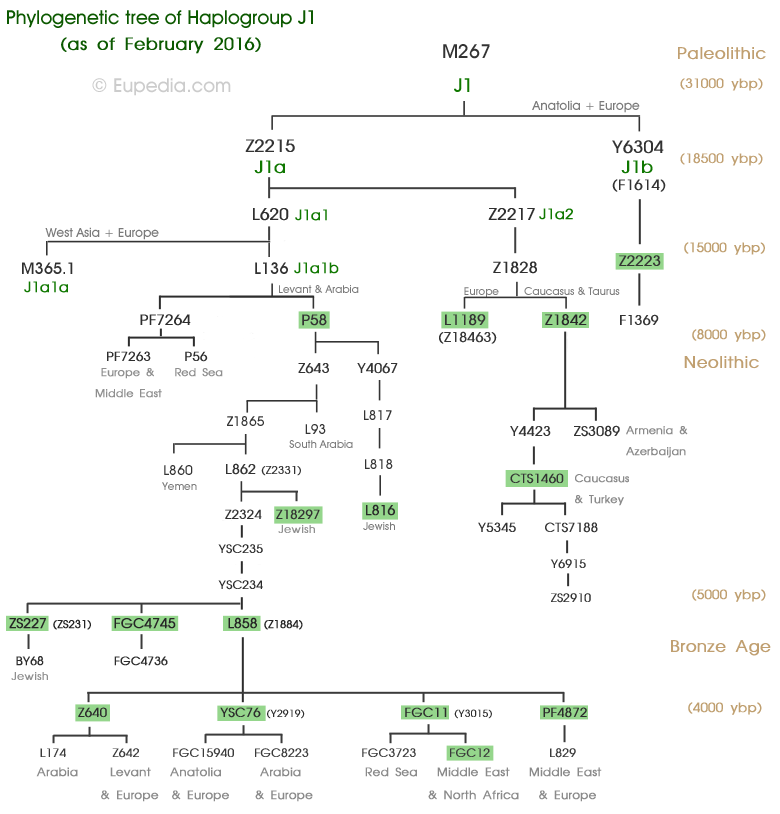 The above phylogenetic tree was is based on the data from Yfull.com, but using the Family Tree DNA nomenclature whenever possible. Main subclades are highlighted in green.
The above phylogenetic tree was is based on the data from Yfull.com, but using the Family Tree DNA nomenclature whenever possible. Main subclades are highlighted in green.
J1 can be divided in two main groups: the very large J1-P58 subclade, and the other branches of J1.
Southwest Asian J1-P58
J1-P58 (J1a2b on the ISOGG tree, formerly known as J1e, then as J1c3) is by far the most widespread subclade of J1. It is a typically Semitic haplogroup, making up most of the population of the Arabian peninsula, where it accounts for approximately 40% to 75% of male lineages.
J1-P58 is thought to have expanded from eastern Anatolia and the southern Caucasus to the Taurus and Zagros mountains, then Mesopotamia, and eventually the Arabian peninsula at the end of the last Ice Age (12,000 years ago) with the seasonal migrations of goat and sheep pastoralists . It is during the Neolithic that subclades like L860 and L93 would have reached the mountainous parts of the southern Arabian peninsula (Yemen, Oman), whereas L816 and L862 remained in the Fertile Crescent.
Arabic speakers recolonised the Arabian peninsula in the Bronze Age from the north-west of the peninsula, close to present-day Jordan. The rise of Islam in the 7th century CE played a major part in the re-expansion of J1 from Arabia throughout the Middle East, as well as to North Africa, and to a lower extent to Sicily, Spain and Portugal.
The numerous subclades downstream of L858 represent the expansion of Semitic languages, such as Akkadian, Ugaritic, Phoenician, Aramaic and Hebrew. Only the FGC12 subclade is linked to the propagation of Islam and the Arabic language from Saudi Arabia from the 7th century CE. Yet even this subclade is shared by the Bedouins and a minority of Jews.
About 20% of Jewish people belong haplogroup J1-P58, most of whom are also positive either for ZS227, L858 (Z642, YSC76 and FGC12) or Z18297. L816 represents a minority of Jewish J1. ZS227 comprises the Cohen Modal Haplotype . In the Hebrew Bible, the common ancestor of all Cohens is identified as Aaron, the brother of Moses. Roughly half of all Cohanim belong to J1-ZS227. The Cohanim haplotype (YCAII=22-22) of ZS227 matches the Z18271 deep clade.
J1-YSC234 (aka Z2313 or Z2329) was found in a pre-Ptolemaic Egyptian mummy dating from c. 660 BCE (Schuenemann et al. (2017)). This clade developped about 5,800 years old, which is about the same age as the first Semitic languages. The mummy is much more recent though, but it proves that J1-YSC234 was present in Egypt long before the Arabic expansion, and therefore that this lineage can be seen as more widely Semitic.
All three branches of J1-L858 (S640, YSC76 and FGC11) are found in Europe , principally in Spain, Italy, central and eastern Europe. Their relatively recent time of divergence with their Middle Eastern cousins (Late Bronze Age to Iron Age) suggests that they would have arrived with the Phoenicians (Sicily, Sardinia, Spain), and later in greater numbers with the Jewish diaspora. Spain and Portugal have the highest percentage of FGC12 in Europe, but this amounts to about 12% of J1 lineages, i.e. less than 0.5% of the population, suggesting that the Arabs had a much smaller genetic impact on the Iberian population than the Jews and the Phoenicians.
Erythrean, Ethiopian, Somali and Sudanese people possess various subclades of J1, but especially J1-P56, which probably arrived during the Neolithic, and the Arabic J1-FGC12, which brought Islam to the region.
STR haplotypes within J1-P58
For men who tested their Y-DNA with companies such as Family Tree DNA, iGenea or Genebase, certain subclades can be easily identified by specific STR markers.
- J1-L93 (aka L92.1) and J1-L386 are both defined by DYS446=13. L93 has a DYS641 value smaller or equal to 10. L386 can be identified by YCAIIa=17. L93 is retricted to Saudi Arabia, Yemen and Oman, while L386 was found chiefly the United Arab Emirates.
- J1-Z640 (aka Z641 or Z644) is particularly common in Syria and Lebanon, but also has been found also in Turkey, Israel and in the Arabian peninsula, as well as well as in many European countries, particularly Spain. It was very probably found among the ancient Phoenicians. Z640+ members typically have DYS561=14. Its subclade L174.1 can be identifed by the STR value DYS594=11.
- J1-L817 is a major Jewish cluster, also defined by L818 and DYS392=13. It has been found a far as China. Most Jewish members belong to its subclade L816.
- J1-L823 (under FGC15940) corresponds to DYS452=30. It is is a Jewish cluster found mostly in central and eastern Europe, but also in Spain.
- J1-L1253 corresponds to DYS557>18. It appears to be limited to Britain and Ireland.
- J1-L1279 corresponds to DYS497=16. A rare subclade identified in the United Arab Emirates.
- J1-L444 is a small Arabic subclade defined by DYS531=12.
- Both L615 (found in the United Arab Emirates) and L859 (found in Iraq and Saudi Arabia) share an STR value for DYS485 around 14.

| Is J1-P58 the main Arabic paternal lineage? |
|---|
|
Looking at the map of J1-P58, it is easy to assume that P58 is a marker of Arabic ancestry because it reaches its maximum frequency in and around the Arabian peninsula. That would be an oversimplification. It is important to make a clear distinction between people who speak Arabic and those who are genetically Arabic. These are two completely different things. For comparison, people who speak languages descended from Latin (French, Spanish, Portuguese, Italian, Romanian) are not necessarily descended from the ancient Romans of the Latium. Even those who do may not have more than a tiny fraction of their genome which was inherited from actual Roman ancestors. This is why most Romance-language speakers today cannot be considered as genetically Roman. Most present-day Arabic speakers outside the Arabian Peninsula are likewise only very partially or not all Arabic genetically. In the northern half of the Middle East, most of the people who call themselves Arabs of today are in fact mainly descendants of other historic peoples, such as the Phoenicians, Assyrians, Babylonians, or even the Hurrians. Most of these peoples are predominantly J2, with many minority haplogroups (E1b1b, G, J1, L, Q, R1a, R1b, T). The confusion comes from the fact that the Arabic language, which appeared a little more than 1,500 years ago, is much more recent than the haplogroup J1 (31,000 years old) or even the P58 subclade (14,000 years old). Even the J1-L858 subclade (5,000 years old), associated with Southwest Asian people, very clearly predates the Arabic language. The common ancestor of the J1-L858 men alive today dates back to approximately 4500 years ago, a time that corresponds to the development of the the oldest Semitic languages, like Akkadian and Amorite. In fact, L858 is not specific to the Arabian peninsula, but is also found among the Jews (especially Z640 subclade), Lebanese, Syrians and Iraqi, among others. In other words L858 cover all the region where ancient Semitic languages were spoken, well before Arabic even existed. That is why L858 should be seen as more widely Semitic and not just Arabic, even if the many Levantines and Mesopotamians were later Arabicised. The Jews are of the these Semitic people who were not Arabicised, and hardly anyone would argue that Jews are Arabs genetically, or vice versa. The true lineage of the historic Arab people (so mainly from Jordan and Saudi Arabia) is J1-FGC12 (aka S21237). This subclade started expanding in the Arabian peninsula a bit over 3,000 years ago and did experience a tremedous expansion in the last 1,300 years, as can be seen in the completely phylogenetic tree on Yfull.com . Nowadays these more genuinely Arabic J1-FGC12 lineages are found throughout the Arabic-speaking world, but they only represent a small minority of lineages in any region but the Arabian peninsula. The other subclades of J1 cannot be considered to be the paternal descendants of first speakers of Arabic. These other J1 lineages were Arabicized alongside other haplogroups (J2, Q1b, etc.) during the Islamic expansion from the 7th century onward. More importantly, J1-FGC12 is not the only haplogroup that spread with the Arabic expansion linked to the diffusion of Islam. Nowadays only 40% of Saudis and 30% of Jordanians belong to J1 (most but not all to FGC12). is another important Arabic lineage, being found in 25% of Jordanians and 10% of Saudis. Like J1-P58, E-M34 it is also shared with their Semitic cousins, the Jews. Haplogroup E1b1b is considered the prime candidate for the origin and dispersal of Afro-Asiatic languages across northern and eastern Africa and south-west Asia. The Semitic languages appear to have originated within a subclade of the M34 branch of E1b1b. One specific deeper subclade is surely associated with the development of Arabic language and with J1-FGC12, but it hasn"t been identified yet. Note that E-M34 itself is many thousands of years old and is also found in non-Semitic countries, including Turkey, Greece, Italy, France and Spain. |
Other subclades
Most of the J1 in the Caucasus, Anatolia and Europe is of the non-J1-P58 variety. These branches diverged from one another during the Ice Age and have nothing to do with Semitic people. In fact the origins of J1 lie around the Caucasus, which explains that all branches other than P58 are most common in that region, as well as in Anatolia and Europe.
J1a1 (M365.1) has been only found in very low frequencies in Western Europe (Western Iberia, French Pyrenees, Belgium and England), in northern Iran and Qatar. It may have come to Europe during the Mesolithic period, or as a very minor Neolithic lineage.
J1a1b1 (P56) is a minor Arabic cluster found on the Red Sea coast of Saudi Arabia, Yemen and Ethiopia. It can be identified by the STR value DYS641=11.
J1a2a (Z1828) is defined by the STR values DYS436=11 and DYS388 L1189 subclade seems to be mostly northern European (+ a few samples in Greece and the Arabian peninsula). Under Z1842 , the CTS1460 subclade is geographically restricted to the Caucasus, Zagros and Taurus, while ZS3089 is found mainly around Armenia and Azerbaijan.
L1189 and Z1842 could have been minor lineages accompanying the main Neolithic haplogroup G2a . That would explain their very wide geographic distribution and low frequency outside their region of origin. Like haplogroup G2a, J1 might have been of the principal lineages to bring domesticated animals to Europe. Both G2a and J1 reach their maximal frequencies in the Caucasus, some ethnic groups being almost exclusively J1 (Kubachis, Kaitaks, Dargins, Avars), while others have extremely high levels of G2a (Shapsugs, North Ossetians). Most of the ethnic groups in the North Caucasus have between 20 and 40% of each haplogroup, which are by far their two dominant haplogroups. In the South Caucasus (Georgia, Armenia, Azerbaijan), haplogroup J2 comes into the admixture and is in fact slightly higher than either J1 or G2a. Armenia stands out of the lot by having a substantial J1-L147.1 (L862) minority (at least one third of all J1, i.e. roughly 4% of the population).
J1b (F1614, Z2223) arose soon after the Last Glacial Maximum (c. 26,500 to 19,000 years before present). It is likely to have spread from the Caucasus to Europe and Anatolia during the Paleolithic and Mesolithic periods. J1-Z2223 matches the STR value DYS446=12. It has been found in Egypt, Anatolia, Italy, Spain, Germany, Belgium and in the British Isles. J1b members negative for Z2223 were found in Finland, Oman and India.









- Серия - Русская фантастика
- Игорь прокопенко - тайны апокалипсиса
- Книга сентября: мемуары Карины Добротворской
- Любовь до смерти и после: «100 писем к Сереже» Карины Добротворской
- Магические способности по дате рождения — особенности расчета
- PR в мифологии Древнего Китая
- Как приготовить мидии Сколько времени занимает варка
- Рулетики из бекона с черносливом и грецким орехом
- Рецепт вареная колбаса с языком Колбасный язык
- Отстранение работника, не прошедшего в установленном порядке обязательный медицинский осмотр Отказаться от медкомиссии на работе
- Табель учета рабочего времени: расшифровка сокращений Как в табеле обозначается учебный оплачиваемый отпуск
- Талисман денежная мельница
- Магия заклинания на латыни с транскрипцией и с переводом
- Гадание на тени от бумаги: правила ворожбы, толкование символов, варианты обрядов
- Характеристика княгини Трубецкой из поэмы Николая Алексеевича Некрасова «Русские женщины
- Все книги про: «жизнь необыкновенные… Краткое содержание жизнь необыкновенных и удивительных
- Что известно о Викторе Золотове и его коррупционных связях
- Россиянам напомнили, как ненасытный "вор и лжец" золотов обкрадывает жителей страны
- Продукты на страже онкологии: полезное и вредное Е вызывающие раковые заболевания
- Крепость орешек Зело жесток сей орех был